Плазматическая мембрана строение и функции кратко. Интегральные белки мембран
Строение и функции биологических мембран
Биологические мембраны– общее название функционально активных поверхностных структур, ограничивающих клетки (клеточные, или плазматические мембраны) и внутриклеточные органеллы (мембраны митохондрий, ядер, лизосом, эндоплазматического ретикулума и др.). Они содержат в своем составе липиды, белки,гетерогенные молекулы (гликопротеины,гликолипиды)и в зависимости от выполняемой функции многочисленные минорные компоненты: коферменты, нуклеиновые кислоты, антиоксиданты, каротиноиды, неорганические ионы и т. п.
Согласованное функционирование мембранных систем – рецепторов, ферментов, транспортных механизмов – помогает поддерживать гомеостаз клетки и в то же время быстро реагировать на изменения внешней среды.
К основным функциям биологических мембран можно отнести:
· отделение клетки от окружающей среды и формирование внутриклеточных компартментов (отсеков);
· контроль и регулирование транспорта огромного многообразия веществ через мембраны;
· участие в обеспечении межклеточных взаимодействий, передаче внутрь клетки сигналов;
· преобразование энергии пищевых органических веществ в энергию химических связей молекул АТФ.
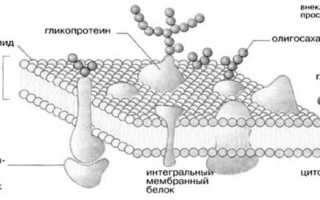
Молекулярная организация плазматической (клеточной) мембраны у всех клеток примерно одинакова: она состоит из двух слоев липидных молекул с множеством включенных в нее специфических белков. Одни мембранные белки обладают ферментативной активностью, тогда как другие связывают питательные вещества из окружающей среды и обеспечивают их перенос в клетку через мембраны. Мембранные белки различают по характеру связи с мембранными структурами. Одни белки, называемые внешними или периферическими, непрочно связаны с поверхностью мембраны, другие, называемые внутренними или интегральными, погружены внутрь мембраны. Периферические белки легко экстрагируются, тогда как интегральные белки могут быть выделены только при помощи детергенов или органических растворителей. На рис. 4 представлена структура плазматической мембраны.
Внешние, или плазматические, мембраны многих клеток, а также мембраны внутриклеточных органелл, например, митохондрий, хлоропластов удалось выделить в свободном виде и изучить их молекулярный состав. Во всех мембранах имеются полярные липиды в количестве, составляющем в зависимости от типа мембран от 20 до 80% ее массы, остальное приходится главным образом на долю белков. Так, в плазматических мембранах животных клеток количество белков и липидов, как правило, примерно одинаково; во внутренней митохондриальной мембране содержится около 80% белков и только 20% липидов, а в миелиновых мембранах клеток мозга наоборот, около 80% липидов и только 20% белков.
Рис. 4. Структура плазматической мембраны
Липидная часть мембран представляет собой смесь различного рода полярных липидов. Полярные липиды, к числу которых относятся фосфоглицеролипиды, сфинголипиды, гликолипиды не запасаются в жировых клетках, а встраиваются в клеточные мембраны, причем в строго определенных соотношениях.
Все полярные липиды в мембранах постоянно обновляются в процессе метаболизма, при нормальных условиях в клетке устанавливается динамическое стационарное состояние, при котором скорость синтеза липидов равна скорости их распада.
В мембранах животных клеток присутствуют в основном фосфоглицеролипиды и в меньшей степени сфинголипиды; триацилглицеролы обнаруживаются лишь в следовых количествах. Некоторые мембраны животных клеток, в особенности наружная плазматическая мембрана, содержит значительные количества холестерола и его эфиров (рис.5).
Рис.5. Мембранные липиды
В настоящее время общепринятой моделью строения мембран является жидкостно-мозаичная, предложенная в 1972 году С. Синджером и Дж. Николсоном.
Согласно ей белки можно уподобить айсбергам, плавающим в липидном море. Как уже указывалось выше, существуют 2 типа мембранных белков: интегральные и периферические. Интегральные белки пронизывают мембрану насквозь, они являются амфипатическими молекулами. Периферические белки не пронизывают мембрану и связаны с ней менее прочно. Основной непрерывной частью мембраны, то есть ее матриксом, служит полярный липидный бислой. При обычной для клетки температуре матрикс находится в жидком состоянии, что обеспечивается определенным соотношением между насыщенными и ненасыщенными жирными кислотами в гидрофобных хвостах полярных липидов.
Жидкостно-мозаичная модель предполагает также, что на поверхности расположенных в мембране интегральных белков имеются R-группы аминокислотных остатков (в основном гидрофобные группы, за счет которых белки как бы «растворяются» в центральной гидрофобной части бислоя). В то же время, на поверхности периферических, или внешних белков, имеются в основном гидрофильные R-группы, которые притягиваются к гидрофильным заряженным полярным головкам липидов за счет электростатических сил. Интегральные белки, а к ним относятся ферменты и транспортные белки, обладают активностью только в том случае, если находятся внутри гидрофобной части бислоя, где они приобретают необходимую для проявления активности пространственную конфигурацию (рис.6). Следует еще раз подчеркнуть, что ни между молекулами в бислое, ни между белками и липидами бислоя не образуется ковалентных связей.
Рис.6. Мембранные белки
Мембранные белки могут свободно перемещаться в латериальной плоскости. Периферические белки буквально плавают на поверхности бислойного «моря», а интегральные белки, подобно айсбергам, почти полностью погружены в углеводородный слой.
В большинстве своем мембраны ассиметричны, то есть имеют неравноценные стороны. Эта ассиметричность проявляется в следующем:
· во-первых, в том, что внутренняя и внешняя стороны плазматических мембран бактериальных и животных клеток различаются по составу полярных липидов. Так, например, внутренний липидный слой мембран эритроцитов человека содержит в основном фосфатидилэтаноламин и фосфатидилсерин, а внешний – фосфатидилхолин и сфингомиелин.
· во-вторых, некоторые транспортные системы в мембранах действуют только в одном направлении. Например, в мембранах эритроцитов имеется транспортная система («насос»), перекачивающая ионы Nа + из клетки в окружающую среду, а ионы К + – внутрь клетки за счет энергии, освобождающейся при гидролизе АТФ.
· в-третьих, на внешней поверхности плазматических мембран содержится очень большое число олигосахаридных группировок, представляющих собой головки гликолипидов и олигосахаридные боковые цепи гликопротеинов, тогда как на внутренней поверхности плазматической мембраны олигосахаридных группировок практически нет.
Ассиметричность биологических мембрам сохраняется за счет того, что перенос индивидуальных молекул фосфолипидов с одной стороны липидного бислоя на другую очень затруднен по энергетическим соображениям. Полярная молекула липида способна свободно перемещаться на своей стороне бислоя, но ограничена в возможности перескочить на другую сторону.
Подвижность липидов зависит от относительного содержания и типа присутствующих ненасыщенных жирных кислот. Углеводородная природа жирнокислотных цепей сообщает мембране свойства текучести, подвижности. В присутствии цис-ненасыщенных жирных кислот силы сцепления между цепями слабее, чем в случае одних насыщенных жирных кислот, и липиды сохраняют высокую подвижность и при низкой температуре.
На внешней стороне мембран имеются специфические распознающие участки, функция которых состоит в распознавании определенных молекулярных сигналов. Например, именно посредством мембраны некоторые бактерии воспринимают незначительные изменения концентрации питательного вещества, что стимулирует их движение к источнику пищи; это явление носит название хемотаксиса.
Мембраны различных клеток и внутриклеточных органелл обладают определенной специфичностью, обусловленной их строением, химическим составом и функциями. Выделяют следующие основные группы мембран у эукариотических организмов:
· плазматическая мембрана (наружная клеточная мембрана, плазмалемма),
· мембраны аппарата Гольджи, митохондрий, хлорпластов, миелиновых оболочек,
У прокариотических организмов помимо плазматической мембраны существуют внутрицитоплазматические мембранные образования, у гетеротрофных прокариот они называются мезосомами. Последние образуются впячиванием внуть наружной клеточной мембраны и в некоторых случаях сохраняют с ней связь.
Мембрана эритроцитовсостоит из белков (50%), липидов (40%) и углеводов (10%). Основная часть углеводов (93%) связана с белками, остальная – с липидами. В мембране липиды расположены асимметрично в отличие от симметричного расположения в мицеллах. Например, кефалин находится преимущественно во внутреннем слое липидов. Такая асимметрия поддерживается, по-видимому, за счет поперечного перемещения фосфолипидов в мембране, осуществляемого с помощью мембранных белков и за счет энергии метаболизма. Во внутреннем слое эритроцитарной мембраны находятся в основном сфингомиелин, фосфатидилэтаноламин, фосфатидилсерин, в наружном слое – фосфатидилхолин. Мембрана эритроцитов содержит интегральный гликопротеин гликофорин, состоящий из 131 аминокислотного остатка и пронизывающий мембрану, и так называемый белок полосы 3, состоящий из 900 аминокислотных остатков. Углеводные компоненты гликофорина выполняют рецепторную функцию для вирусов гриппа, фитогемагглютининов, ряда гормонов. В эритроцитарной мембране обнаружен и другой интегральный белок, содержащий мало углеводов и пронизывающий мембрану. Его называют туннельным белком (компонент а), так как предполагают, что он образует канал для анионов. Периферическим белком, связанным с внутренней стороной эритроцитарной мембраны, является спектрин.
Миелиновые мембраны, окружающие аксоны нейронов, многослойны, в них присутствует большое количество липидов (около 80%, половина из них – фосфолипидов). Белки этих мембран важны для фиксации лежащих друг над другом мембранных солев.
Мембраны хлоропластов. Хлоропласты покрыты двухслойной мембраной. Наружная мембрана имеет некоторое сходство с таковой у митохондрий. Помимо этой поверхностной мембраны в хлоропластах имеется внутренняя мембранная система – ламеллы. Ламеллы образуют или уплощенные пузырьки – тилакоиды, которые, располагаясь друг над другом, собираются в пачки (граны) или формируют мембранную систему стромы (ламеллы стромы). Ламеллы гран и стромы наружной стороне мембраны тилакоидов сосредоточены гидрофильные группировки, галакто- и сульфолипидов. Фитольная часть молекулы хлорофилла погружена в глобулу и находится в контакте в гидрофобными группами белков и липидов. Порфириновые ядра хлорофилла в основном локализованы между соприкасающимися мембранами тилакоидов гран.
Внутренняя (цитоплазматическая) мембрана бактерий по структуре сходна с внутренними мембранами хлоропластов и митохондрий. В ней локализованы ферменты дыхательной цепи, активного транспорта; ферменты, участвующие в образовании компонентов мембраны. Преобладающим компонентом бактериальных мембран являются белки: соотношение белок/липид (по массе) равно 3:1. Наружная мембрана грамотрицательных бактерий по сравнению с цитоплазматической содержит меньшее количество различных фосфолипидов и белков. Обе мембраны различаются по липидному составу. Во внешней мембране находятся белки, образующие поры для проникновения многих низкомолекулярных веществ. Характерным компонентом наружной мембраны является также специфический липополисахарид. Ряд белков наружной мембраны служит рецепторами для фагов.
Мембрана вирусов. Среди вирусов мембранные структуры характерны для содержащих нуклеокапсид, который состоит из белка и нуклеиновой кислоты. Это «ядро» вирусов окружено мембраной (оболочка). Она также состоит из двойного слоя липидов с включенными в него гликопротеинами, расположенными в основном на поверхности мембраны. У ряда вирусов (микровирусы) в мембраны входит 70-80% всех белков, остальные белки содержатся в нуклеокапсиде.
Таким образом, мембраны клеток представляют собой очень сложные структуры; составляющие их молекулярные комплексы образуют упорядоченную двумерную мозаику, что придает поверхности мембран биологическую специфичность.
Плазматическая мембрана: характеристики, строение и функции
Клеточная мембрана, которую также называют плазмалемма, цитолемма или же плазматическая мембрана — является молекулярной структурой, эластичной по своей природе, которая состоит из различных белков и липидов. Она отделяет содержание любой клетки от внешней среды, тем самым регулируя ее защитные свойства, а также обеспечивает обмен между внешней средой и непосредственно внутренним содержимым клетки.
Плазматическая мембрана
Плазмалемма — это перегородка, находящаяся внутри, непосредственно за оболочкой. Она делит клетку на определенные отсеки, которые направлены на компартменты или же органеллы. В них содержатся специализированные условия среды. Клеточная стенка полностью закрывает всю клеточную мембрану. Она выглядит как двойной слой молекул.
Основные сведения
Состав плазмалеммы — это фосфолипиды или же, как их еще называют, сложные липиды. Фосфолипиды имеют несколько частей: хвост и головку. Специалисты называют гидрофобные и гидрофильные части: в зависимости от строения животной или растительной клетки. Участки, которые именуются головкой — обращены внутрь клетки, а хвосты — наружу. Плазмалеммы по структуре являются инвариабельными и очень похожи у различных организмов; чаще всего исключение могут составить археи, у которых перегородки состоят из различных спиртов и глицерина.
Толщина плазмалеммы приблизительно 10 нм.
В малом содержании в состав биологической мембраны входят некоторые виды белков. Например, белки которые пронизывают всю мембрану насквозь, их называют интегральными. Мембраны, которые входят в состав и внешнего, и во внутреннего слоя (слой чаще всего бывает липидным), называются полуинтегральными.
Существуют перегородки, которые находятся на внешней стороне или же снаружи части, вплотную прилегающей к мембране — их называют поверхностными. Некоторые виды белка могут быть своеобразными контактными точками для клеточной мембраны и оболочки. Внутри клетки находится цитоскелет и наружная стенка. Определенные виды интегрального белка могут быть использованы как каналы в ионных транспортных рецепторах (параллельно с нервными окончаниями).
Если использовать электронный микроскоп, то можно получить данные, на основе которых можно построить схему строения всех частей клетки, а также основных составляющих и оболочек. Верхний аппарат будет состоять из трех субсистем:
- комплексное надмембранное включение;
- плазматическая мембрана;
- опорно-сократительный аппарат цитоплазмы, который будет иметь субмембранную часть.
К данному аппарату можно отнести цитоскелет клетки. Цитоплазма с органоидами и ядром называется — ядерный аппарат. Цитоплазматическая или, по-другому, плазматическая клеточная мембрана, находится под клеточной оболочкой.
Слово «мембрана» произошло от латинского слова membrum, которое можно перевести как «кожа» или «оболочка». Термин предложили более 200 лет назад и им чаще называли края клетки, но в период, когда началось использование различного электронного оборудования, установили, что плазматические цитолеммы составляют множество различных элементов оболочки.
Элементы чаще всего структурные, такие как:
Одна из первых гипотез относительно молекулярного состава плазмалеммы была выдвинута в 1940 году научным институтом Великобритании. Уже в 1960 году Уильям Робертс предложил миру гипотезу «Об элементарной мембране». Она предполагала, что все плазмалеммы клетки состоят из определенных частей, по сути, являются сформированными по общему принципу для всех царств организмов.
В начале семидесятых годов XX века было открыто множество данных, на основании которых в 1972 году ученые из Австралии предложили новую мозаично-жидкостную модель строения клеток.
Строение плазматической мембраны
Модель 1972-го года является общепризнанной и по сей день. То есть в современной науке, различные ученые, работающие с оболочкой, опираются на теоретический труд «Строение биологической мембраны жидкостно-мозаичной модели».
Молекулы белков связаны с липидным бислоем и пронизывают всю мембрану полностью — интегральные белки (одно из общепринятых названий — это трансмембранные белки).
Оболочка в составе имеет различные углеводные компоненты, которые будут выглядеть как полисахаридная или сахаридная цепь. Цепь, в свою очередь, будет соединена липидами и белком. Соединенные молекулами белка цепи называются гликопротеинами, а молекулами липидов — гликозидами. Углеводы находятся на внешней стороне мембраны и выполняют функции рецепторов в клетках животного происхождения.
Гликопротеин — представляют собой комплекс надмембранных функций. Его еще называют гликокаликс (от греческих слов глик и каликс, что в переводе означает “сладкий” и “чашка”). Комплекс способствует адгезии клеток.
Функции плазматической мембраны
Барьерная
Помогает отделить внутренние составляющие клеточной массы от тех веществ, которые находятся извне. Предохраняет организм от попадания различных веществ, которые будут являться для него чужеродными, и помогает поддерживать внутриклеточный баланс.
Транспортная
Клетка имеет свой «пассивный транспорт» и использует его для уменьшения расхода энергии. Транспортная функция работает в следующих процессах:
- эндоцитоз;
- экзоцитоз;
- натриевый и калиевый обмен.
На внешней стороне мембраны находится рецептор, на участке которого происходит смешивание гормонов и различных регуляторных молекул.
Пассивный транспорт — процесс, при котором вещество проходит через мембрану, при этом энергия не затрачивается. Иными словами, вещество доставляется из области клетки с высокой концентрацией, в ту сторону, где концентрация будет более низкая.
Существует два вида:
- Простая диффузия — присуща маленьким нейтральным молекулам H2O, CO2 и О2 и некоторыми гидрофобным органическим веществам с низкой молекулярной массой и соответственно без проблем проходят через фосфолипиды мембраны. Эти молекулы могут проникать через мембрану вплоть до того времени, пока градиент концентрации будет стабилен и неизменен.
- Облегченная диффузия — характерна для различных молекул гидрофильного типа. Они также могут проходить через мембрану согласно градиенту концентрации. Однако, процесс будет осуществляться с помощью различных белков, которые будут образовывать специфические каналы ионных соединений в мембране.
Активный транспорт — это перемещение различных составляющих через стенку мембраны в противовес градиенту. Такое перенесение требует значительных затрат энергетических ресурсов в клетке. Чаще всего именно активный транспорт является основным источником потребления энергии.
Выделяют несколько разновидностей активного транспорта при участии белков-переносчиков:
- Натриево-калиевый насос. Получение клеткой необходимых минералов и микроэлементов.
- Эндоцитоз — процесс, при котором происходит захват клеткой твердых частиц (фагоцитоз) или же различных капель любой жидкости (пиноцитоз).
- Экзоцитоз — процесс, при котором происходит выделение из клетки определенных частиц во внешнюю окружающую среду. Процесс является противовесом эндоцитоза.
Термин “эндоцитоз” произошел от греческих слов “энда” (изнутри) и “кетоз” (чаша, вместилище). Процесс характеризует захват внешнего состава клеткой и осуществляется при производстве мембранных пузырьков. Этот термин был предложен в 1965 году профессором цитологии из Бельгии Кристианом Бэйлсом, он изучал поглощение различных веществ клетками млекопитающих, а также фагоцитоз и пиноцитоз.
Фагоцитоз
Происходит при захвате клеткой определенных твердых частиц или же живых клеток. А пиноцитоз — это процесс, при котором капли жидкости захватываются клеткой. Фагоцитоз (от греческих слов “пожиратель” и “вместилище”) — процесс при котором очень маленькие объекты живой природы захватываются и поглощаются, так же как и твердые части различных одноклеточных организмов.
Открытие процесса принадлежит физиологу из России — Вячеславу Ивановичу Мечникову, который определил непосредственно процесс, при этом он проводил различные испытания с морскими звездами и крошечными дафниями.
В основе питания одноклеточных гетеротрофных организмов лежит их способность переваривать, а также захватывать различные частицы.
Мечников описал алгоритм поглощения бактерии амебой и общий принцип фагоцитоза:
- адгезия — прилипание бактерий к мембране клетки;
- поглощение;
- образование пузырька с бактериальной клеткой;
- откупоривание пузырька.
Исходя из этого, процесс фагоцитоза состоит из таких этапов:
- Поглощаемая частица крепится к мембране.
- Окружение поглощаемой частицы мембраной.
- Образование мембранного пузырька (фагосома).
- Открепление мембранного пузырька (фагосомы) во внутреннюю часть клетки.
- Объединение фагосомы и лизосомы (переваривание), а также внутреннее перемещение частиц.
Можно наблюдать полное или частичное переваривание.
В случае частичного переваривания чаще всего образуется остаточное тельце, которое будет находиться внутри клетки некоторое время. Те остатки, которые будут непереварены, изымаются (эвакуируются) из клетки путем экзоцитоза. В процессе эволюции эта функция предрасположенности к фагоцитозу постепенно отделилась и перешла от различных одноклеточных к специализированным клеткам (таким как пищеварительная у кишечнополостных и губок), а после к особым клеткам у млекопитающих и человека.
К фагоцитозу предрасположены лимфоциты и лейкоциты в крови. Сам процесс фагоцитоза нуждается в больших затратах энергии и напрямую объединен с активностью внешней клеточной мембраны и лизосомы, при которых находятся пищеварительные ферменты.
Пиноцитоз
Пиноцитоз — это захват поверхностью клетки какой-либо жидкости, в которой находятся различные вещества. Открытие явления пиноцитоза принадлежит ученому Фицджеральду Льюису. Произошло это событие в 1932 году.
Пиноцитоз — это один из основных механизмов, при котором в клетку попадают высокомолекулярные соединения, например, различные гликопротеины или же растворимые белки. Пиноцитозная активность, в свою очередь, невозможна без физиологического состояния клетки и зависит от ее состава и состава окружающей среды. Самый активный пиноцитоз мы можем наблюдать у амебы.
У человека пиноцитоз наблюдается в клетках кишечника, в сосудах, почечных канальцах, а также в растущих ооцитах. Для того чтобы изобразить процесс пиноцитоза, которой будет осуществляться с помощью лейкоцитов человека, можно сделать выпячивание плазматической мембраны. При этом части будут отшнуровываться и отделяться. Процесс пиноцитоза нуждается в затрате энергии.
Этапы процесса пиноцитоза:
- На наружной клеточной плазмалемме появляются тонкие наросты, которые окружают капли жидкости.
- Этот участок внешней оболочки становится тоньше.
- Образование мембранного пузырька.
- Стенка прорывается (проваливается).
- Пузырек перемещается в цитоплазме и может слиться с различными пузырьками и органоидами.
Экзоцитоз
Термин произошел от греческих слов “экзо” — наружный, внешний и “цитоз” — сосуд, чаша. Процесс заключается в выделении клеточной частью определенных частиц во внешнюю среду. Процесс экзоцитоза является противоположным пиноцитозу.
В процессе экоцитоза из клетки выходят пузырьки внутриклеточной жидкости и переходят на внешнюю мембрану клетки. Содержимое внутри пузырьков может выделяться наружу, а мембрана клетки сливается с оболочкой пузырьков. Таким образом, большинство макромолекулярных соединений будет происходить именно этим способом.
Экзоцитоз выполняет ряд задач:
- доставка молекул на внешнюю клеточную мембрану;
- транспортировка по всей клетке веществ, которые будут нужны для роста и увеличения площади мембраны, например, определенных белков или же фосфолипидов;
- освобождение или соединение различных частей;
- выведение вредных и токсических продуктов, которые появляются при метаболизме, например, соляной кислоты секретируемой клетками слизистой оболочки желудка;
- транспортировка пепсиногена, а также сигнальных молекул, гормонов или нейромедиаторов.
Специфические функции биологических мембран:
- генерация импульса, происходящего на нервном уровне, внутри мембраны нейрона;
- синтез полипептидов, а также липидов и углеводов шероховатой и гладкой сети эндоплазматической сетки;
- изменение световой энергии и ее преобразование в энергию химическую.
Видео
Из нашего видео вы узнаете много интересного и полезного о строении клетки.
Клеточная мембрана
Клеточная мембрана также называется плазматической (или цитоплазматической) мембраной и плазмалеммой. Данная структура не только отделяет внутреннее содержимое клетки от внешней среды, но также входит с состав большинства клеточных органелл и ядра, в свою очередь отделяя их от гиалоплазмы (цитозоля) — вязко-жидкой части цитоплазмы. Договоримся называть цитоплазматической мембраной ту, которая отделяет содержимое клетки от внешней среды. Остальными терминами обозначать все мембраны.
Строение клеточной мембраны
В основе строения клеточной (биологической) мембраны лежит двойной слой липидов (жиров). Формирование такого слоя связано с особенностями их молекул. Липиды не растворяются в воде, а по-своему в ней конденсируются. Одна часть отдельно взятой молекулы липида представляет собой полярную головку (она притягивается водой, т. е. гидрофильна), а другая — пару длинных неполярных хвостов (эта часть молекулы отталкивается от воды, т. е. гидрофобна). Такое строение молекул заставляет их «прятать» хвосты от воды и поворачивать к воде свои полярные головки.
В результате образуется двойной липидный слой, в котором неполярные хвосты находятся внутри (обращены друг к другу), а полярные головки обращены наружу (к внешней среде и цитоплазме). Поверхность такой мембраны гидрофильна, а внутри она гидрофобна.
В клеточных мембранах среди липидов преобладают фосфолипиды (относятся к сложным липидам). Их головки содержат остаток фосфорной кислоты. Кроме фосфолипидов есть гликолипиды (липиды + углеводы) и холестерол (относится к стеролам). Последний придает мембране жесткость, размещаясь в ее толще между хвостами остальных липидов (холестерол полностью гидрофобный).
За счет электростатического взаимодействия, к заряженным головкам липидов присоединяются некоторые молекулы белков, которые становятся поверхностными мембранными белками. Другие белки взаимодействуют с неполярными хвостами, частично погружаются в двойной слой или пронизывают его насквозь.
Таким образом, клеточная мембрана состоит из двойного слоя липидов, поверхностных (периферических), погруженных (полуинтегральных) и пронизывающих (интегральных) белков. Кроме того, некоторые белки и липиды с внешней стороны мембраны связаны с углеводными цепями.
Это жидкостно-мозаичная модель строения мембраны была выдвинута в 70-х годах XX века. До этого предполагалась бутербродная модель строения, согласно которой липидный бислой находится внутри, а с внутренней и наружной стороны мембрана покрыта сплошными слоями поверхностных белков. Однако накопление экспериментальных данных опровергло эту гипотезу.
Толщина мембран у разных клеток составляет около 8 нм. Мембраны (даже разные стороны одной) отличаются между собой по процентному соотношению различных видов липидов, белков, ферментативной активности и др. Какие-то мембраны более жидкие и более проницаемые, другие более плотные.
Разрывы клеточной мембраны легко сливаются из-за физико-химических особенностей липидного бислоя. В плоскости мембраны липиды и белки (если только они не закреплены цитоскелетом) перемещаются.
Функции клеточной мембраны
Большинство погруженных в клеточную мембрану белков выполняют ферментативную функцию (являются ферментами). Часто (особенно в мембранах органоидов клетки) ферменты располагаются в определенной последовательности так, что продукты реакции, катализируемые одним ферментом, переходят ко второму, затем третьему и т. д. Образуется конвейер, который стабилизируют поверхностные белки, т. к. не дают ферментам плавать вдоль липидного бислоя.
Клеточная мембрана выполняет отграничивающую (барьерную) от окружающей среды и в то же время транспортную функции. Можно сказать, это ее самое главное назначение. Цитоплазматическая мембрана, обладая прочностью и избирательной проницаемостью, поддерживает постоянство внутреннего состава клетки (ее гомеостаз и целостность).
При этом транспорт веществ происходит различными способами. Транспорт по градиенту концентрации предполагает передвижение веществ из области с их большей концентрацией в область с меньшей (диффузия). Так, например, диффундируют газы (CO2, O2).
Бывает также транспорт против градиента концентрации, но с затратой энергии.
Транспорт бывает пассивным и облегченным (когда ему помогает какой-нибудь переносчик). Пассивная диффузия через клеточную мембрану возможна для жирорастворимых веществ.
Есть особые белки, делающие мембраны проницаемыми для сахаров и других водорастворимых веществ. Такие переносчики соединяются с транспортируемыми молекулами и протаскивают их через мембрану. Так переносится глюкоза внутрь эритроцитов.
Пронизывающие белки, объединяясь, могут образовывать пору для перемещения некоторых веществ через мембрану. Такие переносчики не перемещаются, а образуют в мембране канал и работают аналогично ферментам, связывая определенное вещество. Перенос осуществляется благодаря изменению конформации белка, благодаря чему в мембране образуются каналы. Пример — натрий-калиевый насос.
Транспортная функция клеточной мембраны эукариот также реализуется за счет эндоцитоза (и экзоцитоза). Благодаря этим механизмам в клетку (и из нее) попадают крупные молекулы биополимеров, даже целые клетки. Эндо- и экзоцитоз характерны не для всех клеток эукариот (у прокариот его вообще нет). Так эндоцитоз наблюдается у простейших и низших беспозвоночны; у млекопитающих лейкоциты и макрофаги поглощают вредные вещества и бактерии, т. е. эндоцитоз выполняет защитную функцию для организма.
Эндоцитоз делится на фагоцитоз (цитоплазма обволакивает крупные частицы) и пиноцитоз (захват капелек жидкости с растворенными в ней веществами). Механизм этих процессов приблизительно одинаков. Поглощаемые вещества на поверхности клеток окружаются мембраной. Образуется пузырек (фагоцитарный или пиноцитарный), который затем перемещается внутрь клетки.
Экзоцитоз — это выведение цитоплазматической мембраной веществ из клетки (гормонов, полисахаридов, белков, жиров и др.). Данные вещества заключаются в мембранные пузырьки, которые подходят к клеточной мембране. Обе мембраны сливаются и содержимое оказывается за пределами клетки.
Цитоплазматическая мембрана выполняет рецепторную функцию. Для этого на ее внешней стороне располагаются структуры, способные распознавать химический или физический раздражитель. Часть пронизывающих плазмалемму белков с наружней стороны соединены с полисахаридными цепочками (образуя гликопротеиды). Это своеобразные молекулярные рецепторы, улавливающие гормоны. Когда конкретный гормон связывается со своим рецептором, то изменяет его структуру. Это в свою очередь запускает механизм клеточного ответа. При этом могут открываться каналы, и в клетку могут начать поступать определенные вещества или выводиться из нее.
Рецепторная функция клеточных мембран хорошо изучена на основе действия гормона инсулина. При связывании инсулина с его рецептором-гликопротеидом происходит активация каталитической внутриклеточной части этого белка (фермента аденилатциклазы). Фермент синтезирует из АТФ циклическую АМФ. Уже она активирует или подавляет различные ферменты клеточного метаболизма.
Рецепторная функция цитоплазматической мембраны также включает распознавание соседних однотипных клеток. Такие клетки прикрепляются друг к другу различными межклеточными контактами.
В тканях с помощью межклеточных контактов клетки могут обмениваться между собой информацией с помощью специально синтезируемых низкомолекулярных веществ. Одним из примеров подобного взаимодействия является контактное торможение, когда клетки прекращают рост, получив информацию, что свободное пространство занято.
Межклеточные контакты бывают простыми (мембраны разных клеток прилегают друг к другу), замковыми (впячивания мембраны одной клетки в другую), десмосомы (когда мембраны соединены пучками поперечных волокон, проникающих в цитоплазму). Кроме того, есть вариант межклеточных контактов за счет медиаторов (посредников) — синапсы. В них сигнал передается не только химическим, но и электрическим способом. Синапсами передаются сигналы между нервными клетками, а также от нервных к мышечным.
Источники:
http://studopedia.ru/9_189522_stroenie-i-funktsii-biologicheskih-membran.html
http://liveposts.ru/articles/education-articles/biologiya/plazmaticheskaya-membrana-harakteristiki-stroenie-i-funktsii
http://biology.su/cytology/cell-membrane